Effects of chilling stress & responses of plants to chilling
Effects of chilling stress & responses of plants to chilling
2. Macroscopic effects of chilling on sensitive species: chilling injuries
- As in the case of other stresses, the severity of the symptoms observed will depend on the severity of the stress applied. In the case of cotton germinating seeds, the earlier manifestation of chilling stress is a slowing down of the germination process.
- More symptoms include;
- stunting,
- the appearance of necrotic areas,
- surface pitting
- external discoloration.
- Banana fruits exposed to severe chilling when green will turn black.
- Other symptoms include rubbery texture and irregular ripening, lignification, woolliness and internal breakdown.
3. Effects of chilling on cellular ultrastructures
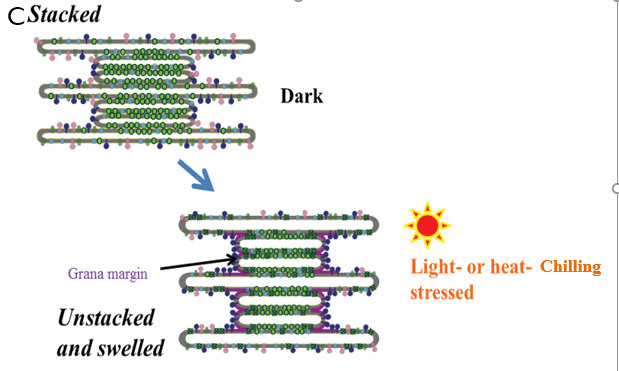
- The subcellular compartment that is mainly affected by chilling is the chloroplast, where the first manifestations of injury are a distortion and swelling of thylakoids as well as a reduction in the size and number of starch granules.
- Thylakoid dilation appears to be related to photo-oxidative conditions produced during chilling in the light.
- In addition chilling will induce the formation of vesicles from the envelope, forming the so-called peripheral reticulum.
- If chilling is prolonged, more severe modifications involving unstacking of grana and eventually disappearance of the chloroplast envelope have been observed.
- In a few other cases, other morphological changes have been observed in the
- nucleus (condensation of chromatin)
- alterations in the appearance of the nucleolus, and in the Golgi apparatus and endoplasmic reticulum, which became dilated.
4. Effects of chilling on biochemical and molecular processes
Lowering temperatures will thermodynamically reduce the kinetics of metabolic reactions.
- Exposure to low temperatures will directly affect the stability and solubility of many proteins or protein complexes.
- Both the effects of chilling on enzymatic rates and on protein stability will affect enzymes involved in metabolism.
- Effects of chilling on photosynthesis-related processes
- During photosynthesis, CO2 combines with ribulose-1,5-bisphosphate to produce glycerate-3-phosphate, which is reduced into triose-phosphates.
- Most triose-phosphate is retained in the chloroplast, with the remainder mainly being exported to the cytosol and converted into sucrose .
- The final conversion steps to sucrose release inorganic phosphate (Pi) that returns to the chloroplast for ATP synthesis.
- Exposure to low temperatures results in a decrease in triose-phosphate utilization that leads to the accumulation of phosphorylated metabolites, which in turn leads to a decrease in inorganic phosphate (Pi) availability in chloroplasts.
- This Pi status of the plastids in turn reduces the availability of the electron acceptor glycerate-1,3-bisphosphate in the photosynthetic reduction cycle (Calvin cycle). This is the Pi-limitation of photosynthesis observed at the early stage of chilling.
- This imbalance in photostasis favours the accumulation of ROS such as singlet oxygen, hydroxyl radical or hydrogen peroxide, since the normal electron acceptor is not available.
- This ROS accumulation is further amplified by the fact that the activities of the scavenging enzymes are thermodynamically lowered by low temperatures.
- The accumulation of ROS will have deleterious effects. It will lead to peroxidation of membrane lipids.
- Membrane integrity will be disrupted and this will result in ion leakage.
- Lowering the temperature will also cause membranes to be more rigid, thereby leading to a disturbance of all membrane processes (e.g. the opening of ion channels, membrane-associated electron transfer reactions etc.).
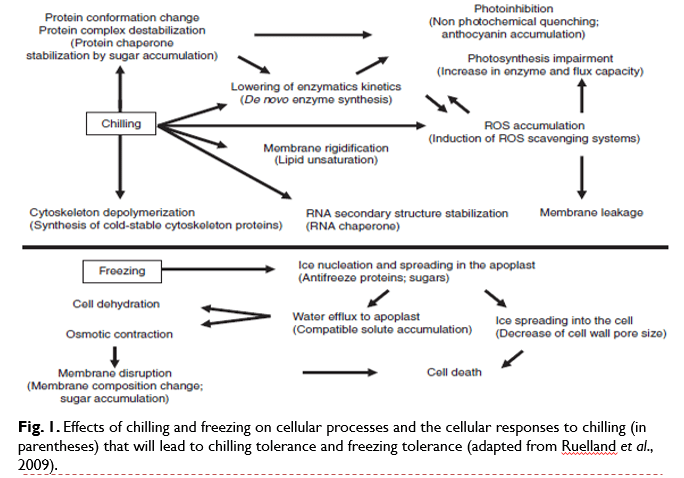
- In addition, the low temperatures will favour the formation of secondary structures in RNA, which can disturb translational processes (Fig. 1).
The Responses to Chilling Temperatures
- Chilling induces a plethora of responses in plants. Some of these responses will participate in the acquisition of chilling tolerance and, in some species, in freezing tolerance.
- This is the molecular explanation of cold hardening or cold-acclimation, i.e. the fact that an exposure to chilling temperatures may, in some species, result in an improved freezing tolerance.
- Stress-related proteins
- Dehydrins
- One of the best-documented responses of plants to chilling treatments is the accumulation of hydrophilic proteins.
- Many of the genes encoding these proteins were first characterized as being responsive to cold, drought and/or abscisic acid (ABA).
- Most of these proteins have therefore been named COR (cold-responsive), LTI (low temperature- induced), RAB (responsive to abscisic acid), KIN (cold-induced) or ERD (early responsive to dehydration).
- They include dehydrins, which define group II of Late Embryogenesis Abundant (LEA) proteins.
- Dehydrins may have a role in freezing tolerance, possibly by preventing the membrane destabilization.
- Some dehydrins also possess cryoprotective properties, have antifreeze activity or act as oxygen radical scavengers
Chilling-induced proteins with antifreeze properties
- Antifreeze proteins (AFPs) prevent ice nucleation and ice crystal formation.
- Plants have no constitutive AFP activity. It is induced by chilling, and clearly plays a role in freezing tolerance.
- AFPs would act by directly interacting with ice in planta and by reducing freezing injury by slowing the growth and recrystallization of ice.
Cold-shock proteins and RNA-binding proteins
- One of the major problems that living organisms have to deal with when exposed to low temperatures is the formation and stabilization of RNA secondary structures.
- The protective proteins that has been named cold shock proteins (CSP) in bacteria and plant.
- They are suggested to act as RNA chaperones during cold shock inhibiting the formation of RNA secondary structures, which are unfavourable for transcription and translation.
- These proteins have glycine-rich regions. Such proteins exist in plants and are induced by chilling temperatures.
2. Membrane composition changes
Leaves from plants injured by chilling show
- Inhibition of photosynthesis,
- Slower carbohydrate translocation
- Lower respiration rates,
- Inhibition of protein synthesis
- Increased degradation of existing proteins.
- All of these responses appear to depend on a common primary mechanism involving loss of membrane function during chilling.
- For instance, solutes leak from the leaves of chilling sensitive Passiflora maliformis (conch apple; passion fruit) floated on water at 0°C, but not from those of chilling-resistant Passiflora caerulea (passion flower). Loss of solutes to the water reflects damage to the plasma membrane and tonoplast.
- In turn, inhibition of photosynthesis and of respiration reflects injury to chloroplast and mitochondrial membranes.
Why are membranes affected by chilling?
- The lipids in the bilayer have a high percentage of saturated fatty acid chains, and membranes with this composition tend to solidify into semicrystalline state at a temperature well above 0°C.
- As the membranes become less fluid, their protein components can no longer function normally.
- Membrane lipids from chilling-resistant plants often have a greater proportion of unsaturated fatty acids than those from chilling-sensitive plants.
During Acclimation to cool temperatures the activity of desaturase enzymes increases and the proportion of unsaturated lipids rises
This modification allows membranes to remain fluid at lower temperatures.
Fatty acid composition of mitochondria isolated from chilling-resistant and chilling-sensitive species
| Chilling-resistant species | Chilling-sensitive species | |||||
| Cauliflower bud | Turnip root | Pea shoot | Bean shoot | Sweet potato | Maize shoot | |
| Ratio of unsaturated to saturated
fatty acids |
3.2 | 3.9 | 3.8 | 2.8 | 1.7 | 2.1
|
3. Sugars accumulation during an exposure to chilling
- Leaf soluble sugars, including sucrose and raffinose, and of sugar alcohols such as galactinol, increases dramatically during chilling exposure.
- This increase mainly proceeds from the starch degradation pathway.
- During freezing, ice is initially formed in the apoplastic space. The water potential in this compartment therefore decreases. This leads to an exit of water from the cell into the extracellular compartment, resulting in cell dehydration (Fig. 1).
- To avoid this cell dehydration it is necessary to decrease the water potential of the cell.
- The accumulation of carbohydrates within the cell contributes to this by diminishing the difference in water potentials between the apoplastic space and the solution within the cell.
- Second, sugars may be active cryoprotectants towards membranes. The water associated with membranes is required to create the hydrophilic environment necessary to stabilize lipids in a bilayer.
- During freeze-induced dehydration, non-reducing sugars, such as sucrose or trehalose, can replace the lost water in creating this hydrophilic environment.
- Finally, sugars can also have a role in protecting integral membrane protein complexes.
4. Compatible solute other than sugars
- In addition to soluble sugars, compatible solutes are a heterogeneous group of molecules comprising amino acids (Pro, Ala, Gly, Ser) and polyamines.
- It is for instance well documented that proline robustly accumulates under chilling stress.
- Polyamines are organic compounds having two or more primary amino groups. In Arabidopsis submitted for 14 days to a temperature of 4°C, an increase in putrescine, ornithine and citrulline, the precursors of polyamines, have been observed.
- Physiologically, compatible solutes have no adverse metabolic effects even at very high concentrations.
- As already mentioned for sugars, they may act by decreasing the water potential, thus preventing excessive cell dehydration, counter balancing the osmotic effect of ice formation in the apoplastic space.
- They may also stabilize proteins, assist refolding of peptides and stabilize membranes.
5. Photosynthesis and photosynthesis-related pigments
- Cold temperatures will lead to an early imbalance between the light energy harvested by antennae and its use by biochemical processes.
- In the long term, plants have evolved diverse strategies to cope with low temperatures.
- In evergreen conifers such as Pinus sylvestris, cold acclimation induces cessation of primary growth and a decrease in the sink demand for photoassimilates. The cessation of growth decreases the capacity for energy utilization.
- To attain photostasis under these conditions, over-wintering conifers exhibit long-term changes in the organization of the photosynthetic apparatus, which includes a decrease in the number of functional PS II reaction centers, a loss of light-harvesting chlorophylls and the formation of a large thylakoid/protein aggregate involving the light-harvesting complex II and PS I and PS II.
- This aggregation is associated with an increased formation of zeaxanthin, through the xanthophyll cycle.
- It is the cycle by which violaxanthin is rapidly and reversibly de-epoxidized into zeaxanthin via the intermediate antheraxanthin.
- Antheraxanthin and zeaxanthin are quenching pigments, unable to pass their excitation to chlorophyll a.
- This is a protective mechanism that lowers the delivery of energy to PS II. It is a part of non-photochemical quenching (NPQ).
- The formation of the large winter-induced aggregated state is fully reversible upon warming in the spring, with the result that photosynthesis is rapidly resumed without the immediate de novo synthesis of chlorophyll
6. Reactive oxygen species scavenging systems
Reactive oxygen species scavenging systems
- Cold exposure induces an oxidative stress. At low temperature, the enzymatic systems that normally detoxicate ROS will be less efficient.
- In plants subjected to low temperatures, scavenging systems will be induced and activated in response to ROS production.
- Catalase and superoxide dismutase are induced by cold.
- In Arabidopsis thaliana, an increase in ascorbate was observed and was still observed after a 14-day exposure at 4°C.
- Ascorbate is an abundant antioxidant in plants.
- The detoxification of hydrogen peroxide produced in the chloroplasts relies exclusively on the activity of ascorbate peroxidase bound to thylakoid membranes in the vicinity of PS I.
- A correlation can be made between chilling resistance and activation of ROS scavenging systems.